Ferredoxin/Ferredoxin-NAD(P)+ reductase
光合成生物のNAD(P)+光還元系と, 従属栄養生物のferredoxin/flavodoxinを中心とした代謝系
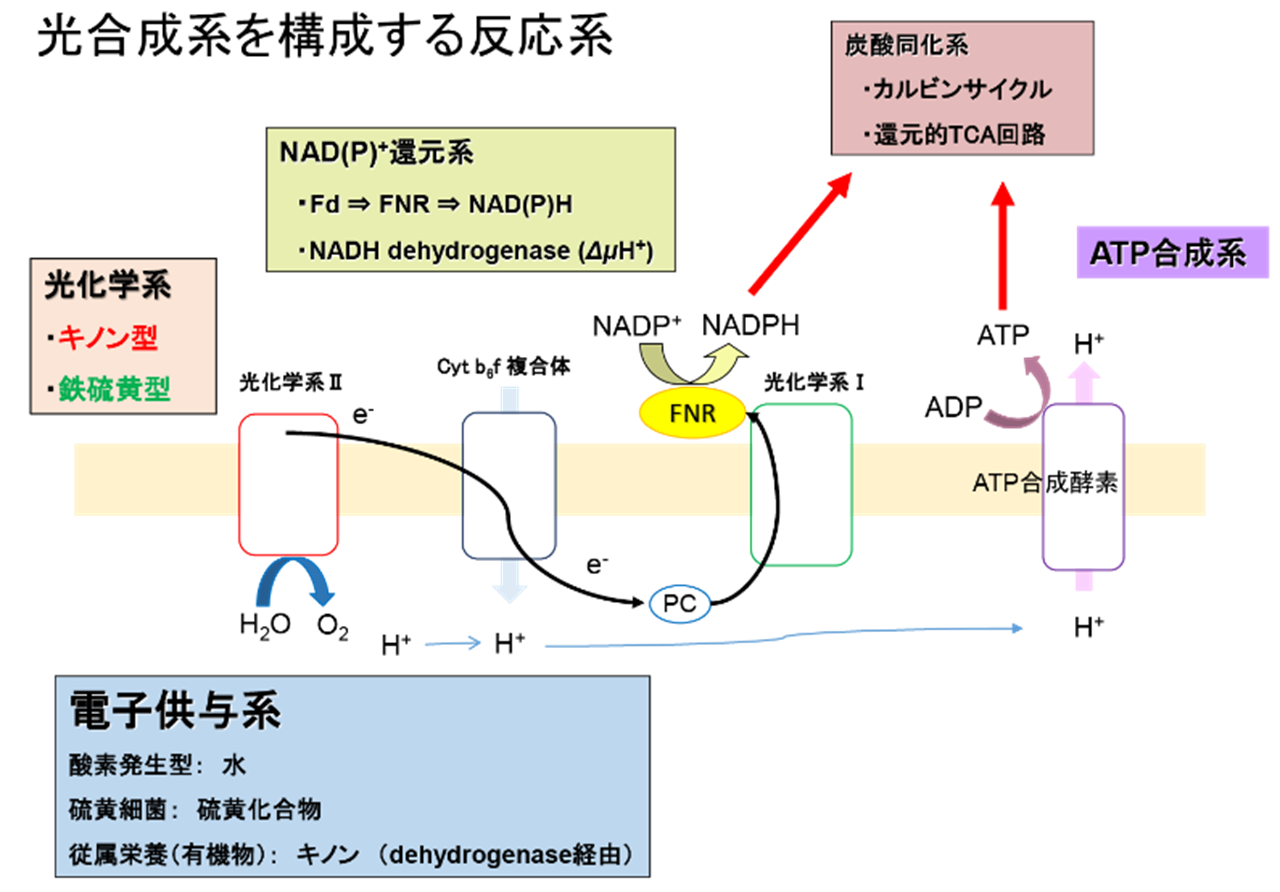
光合成は太陽光エネルギーを利用した同化過程で、電子供与系、光化学系、炭酸同化系などの代謝系が統合されたシステムであり、水以外にも硫化物を利用する電子供与系や、還元的TCA回路を用いる炭酸同化系など、多様な形態が存在します(右図)。光化学系を除いた電子供与系や炭酸同化系は、古細菌や嫌気性細菌などの非光合成生物にも相同な系が存在するため、生物における代謝系の多様性の理解は、生物のエネルギー獲得方法の誕生と進化の過程の理解や、生体エネルギーを利用した問題解決への糸口を与えることが期待されます。私たちは、嫌気性光合成細菌である緑色硫黄細菌の電子供与系からNAD(P)+光還元系までを構成する電子伝達タンパク質(可溶性シトクロム、光化学反応中心、鉄硫黄蛋白質、フラボ蛋白質)の生化学的・物理化学的手法を用いた研究を基に、現在は光合成生物と従属栄養生物由来のferredoxin-NAD(P)+酸化還元酵素(FNR)とその関連酵素・電子伝達タンパク質の研究を、組換え型タンパク質の大量発現、変異導入をはじめとする遺伝子工学的手法、閃光分光法やストップトフロー法などの速度論的手法に加えて、共同研究によるX線結晶構造解析や高磁場NMRなどの構造学的手法を用いて進めています。
1. 細菌由来Ferredoxin (Fd), Ferredoxin-NAD(P)+酸化還元酵素 (FNR)の研究
.png)
FNRはNAD(P)HとFd間の酸化還元反応を触媒するフラボ酵素です。私たちは緑色硫黄細菌C. tepidumの菌体からFNRの単離と遺伝子の帰属に世界に先駆けて成功し、同細菌が植物と同様に鉄硫黄タンパク質とフラボ酵素の構成でNAD(P)+光還元を行うことを明らかにしました。単離したFNRは既知FNRとの相同性が低い新規なFNRで、FNRの単離・精製に関する報告のなかったグラム陽性菌や古細菌にホモログが存在します。様々な細菌由来(緑色硫黄細菌、紅色非硫黄細菌、枯草菌など)のFdとFNRの大量発現系を構築し、大阪大学蛋白質研究所・兵庫県立大学との共同研究により3種のFNRの結晶構造を明らかにし、現在は構造‐機能相関の観点から、基質結合に伴う構造ダイナミクスの変化が反応へ及ぼす影響を明らかにすることを目指して、部位特異的変異体を用いたストップトフロー法による速度論的研究、中間体の構造情報の取得を目指した高磁場NMRや酵素基質複合体のX線結晶構造解析(大阪大学・香川大学との共同研究)を実施しています。
2. Ferredoxin, Flavodoxin, Rubredoxin依存型酵素の研究
生物は嫌気条件下においてFd依存型の酸化還元酵素を用い、炭酸同化、窒素固定、芳香族化合物の分解など様々な反応を行います。嫌気下では好気下と比較して強い還元力を用いることができるため、ユニークかつ有用な反応が多く存在しますが、反応を担う一連の酵素には好気下で活性を失うものが多く、基質特異性や反応機構などに関する研究は進んでいません。特に鉄硫黄クラスターを持つ酵素には有用なものが多いのですが、酸素に弱い酵素が多くみられます。私たちは、これらの依存型酸化還元酵素の単離・精製とその生化学的な性質を明らかにすること、また未知のFd依存型酵素の探索も行っています。
3. 緑色硫黄細菌Chlorobaculum tepidumの光化学反応中心と周辺電子伝達系の研究
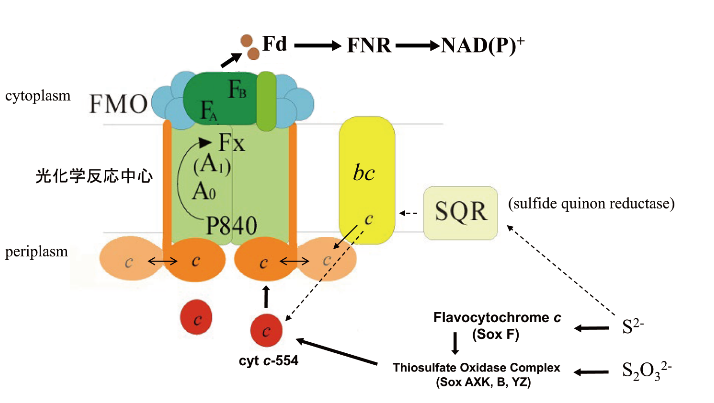
緑色硫黄細菌C. tepidumの光化学反応中心は鉄硫黄型の反応中心で、ferredoxinを直接還元できます。窒素雰囲気の嫌気下で高いFd還元活性を有する反応中心の精製に成功し、フランスとの共同研究による多連閃光を用いた分光測定により、初発電子供与体P840+のシトクロムcによる還元速度と酸化還元平衡から2つの等価なヘムが反応中心当たり2個結合していること、末端電子受容体は3つの鉄硫黄クラスターで、光化学系Iと異なりキノンが電子伝達に関与する可能性が低いことなどを明らかにしました。また神奈川大学理学部との共同研究により、緑色硫黄細菌C. tepidumから可溶性ヘム蛋白質の単離と遺伝子への帰属を行い、無機硫黄化合物から反応中心へ至る電子伝達系の構成因子を明らかにし、個々の電子伝達タンパク質に関する研究を進めています。